Transposons: the ‘jumping genes’ revolutionizing genetics
Written by Anina Werner · 07. January 2025

When Barbara McClintock studied patterns in corn kernels in the 1940s, she made a discovery that would challenge our understanding of genetics. She observed elements that could 'jump' from one location to another in the genome, causing changes in kernel coloration. The identification of these mobile elements – now known as transposons, transposable elements (TEs) or 'jumping genes' – was so revolutionary that it took decades for the scientific community to fully appreciate their significance and adapt them into modern tools for basic research and gene therapy.
McClintock's pioneering work eventually earned her the 1983 Nobel Prize in Physiology or Medicine, recognizing the profound impact of her discovery. Today, we know that transposons exist in various forms, each with unique characteristics. This blog will explain the differences between them, and will explore their roles in evolution, health and genetic engineering applications.
Table of contents
What are transposons?
Transposons are DNA sequences that can move from one location to another within a genome. They are widespread in both prokaryotes and eukaryotes, often making up significant portions of an organism's genetic material. In humans, for instance, transposons account for approximately 50 % of the genome, whereas in corn, they account for up to 90 %.
Types of transposons
Transposons can be grouped into distinct types based on 2 main criteria: their mechanism of movement and their independence. They are classified as retrotransposons or DNA transposons, depending on how they move within the genome, and as autonomous or non-autonomous, based on whether they require other transposons for their mobility.
Retrotransposons: class I TEs
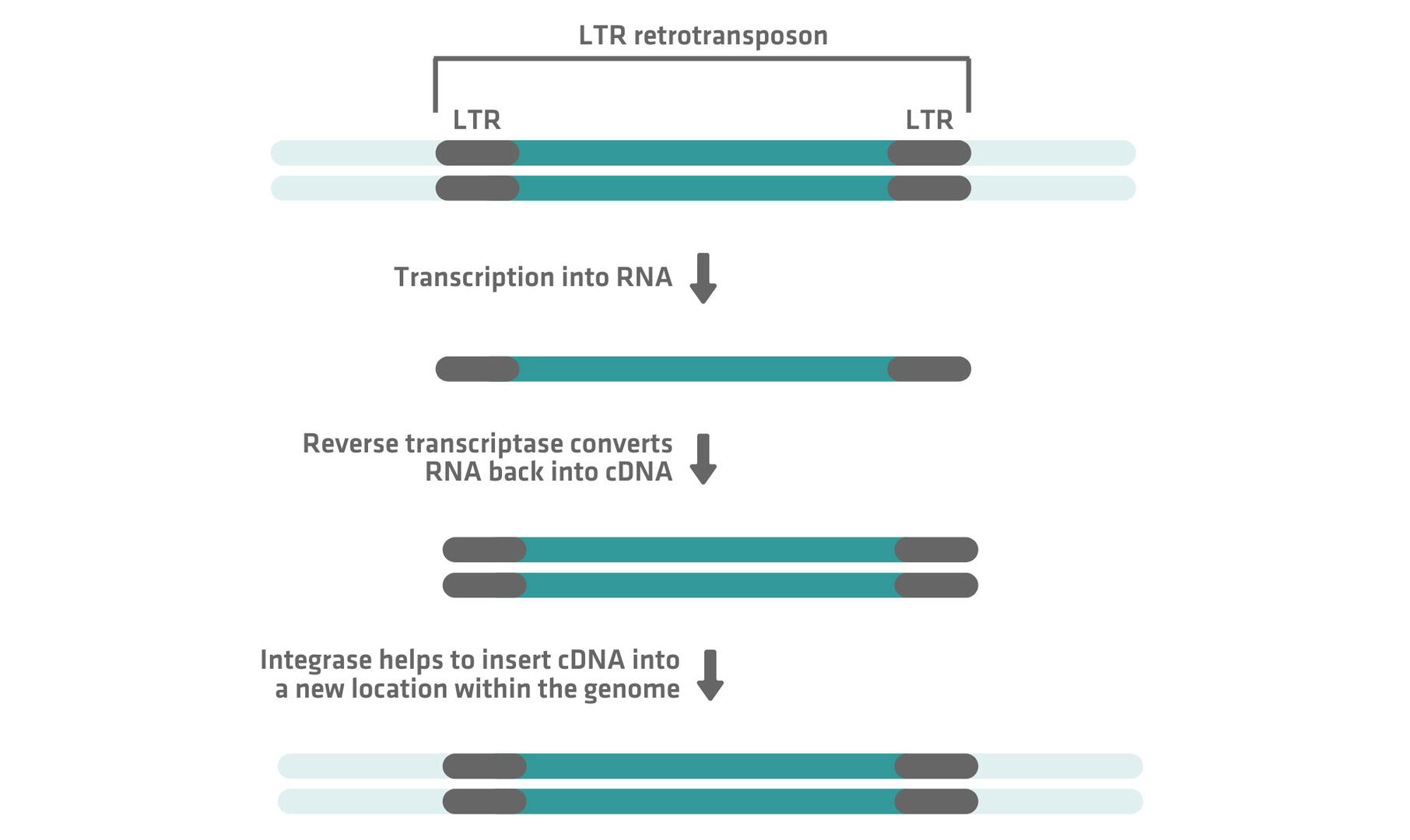
Retrotransposons, also known as class I TEs, use a 'copy-and-paste' mechanism to replicate and insert themselves into new locations within a genome. This process, which involves an RNA intermediate, has 3 key steps and allows retrotransposons to preserve their original copy while creating new insertions elsewhere.
- Transcription: The retrotransposon sequence is first transcribed into RNA using the cell's transcription machinery.
- Reverse transcription: The RNA is then converted into complementary DNA (cDNA).
- Integration: This newly synthesized cDNA is inserted into a different location within the genome.
Retrotransposons can be categorized into 2 main subtypes: LTR and non-LTR retrotransposons. These subtypes are distinguished primarily by their structural characteristics and integration mechanisms.
LTR retrotransposons feature long terminal repeats (LTRs) on both ends of their sequence, and rely on reverse transcriptase and integrase enzymes to integrate into a new genomic location after RNA transcription:
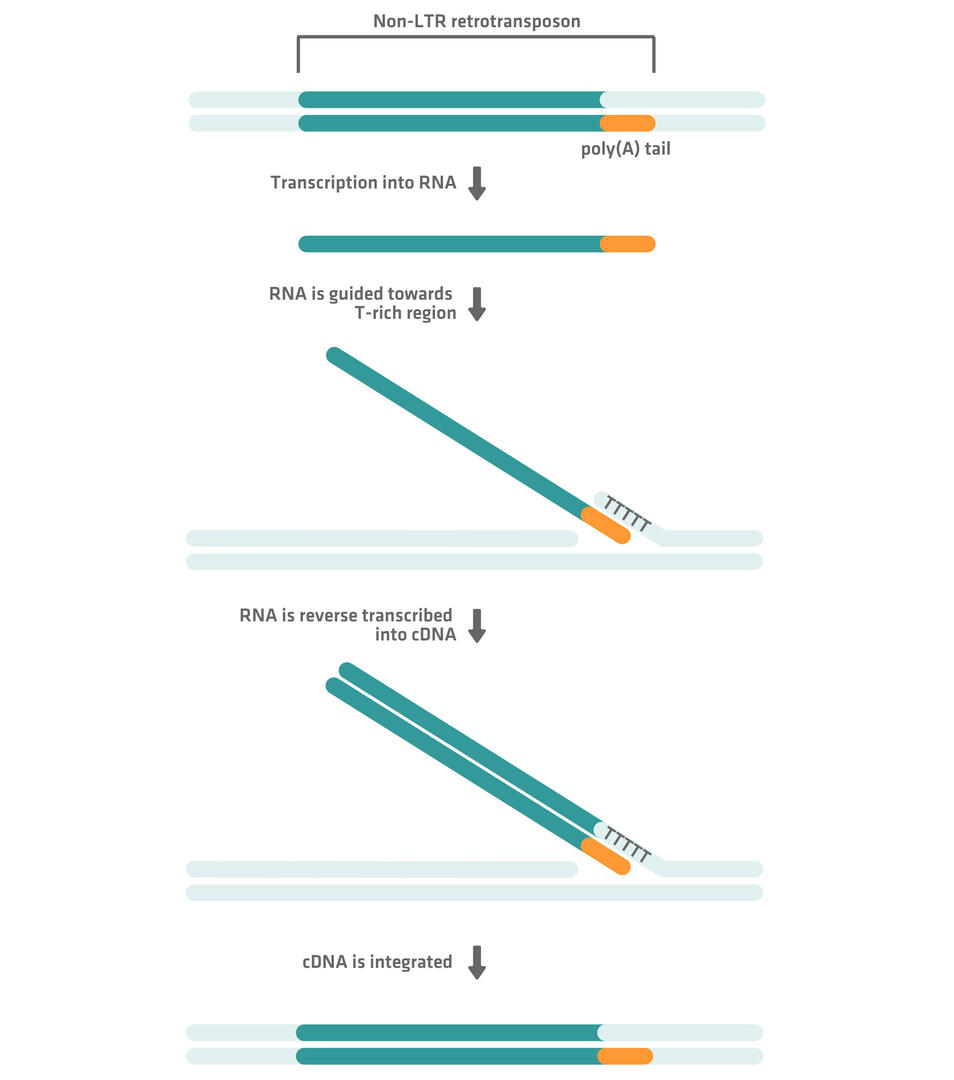
Non-LTR retrotransposons have a poly(A) tail at one end rather than LTRs. The retrotransposon is first transcribed into RNA, which is then guided towards a T-rich region in the genome by an RNA binding protein and another protein with endonuclease and reverse transcriptase activity. The RNA is then reverse transcribed into cDNA and integrated into the genome at the T-rich region:
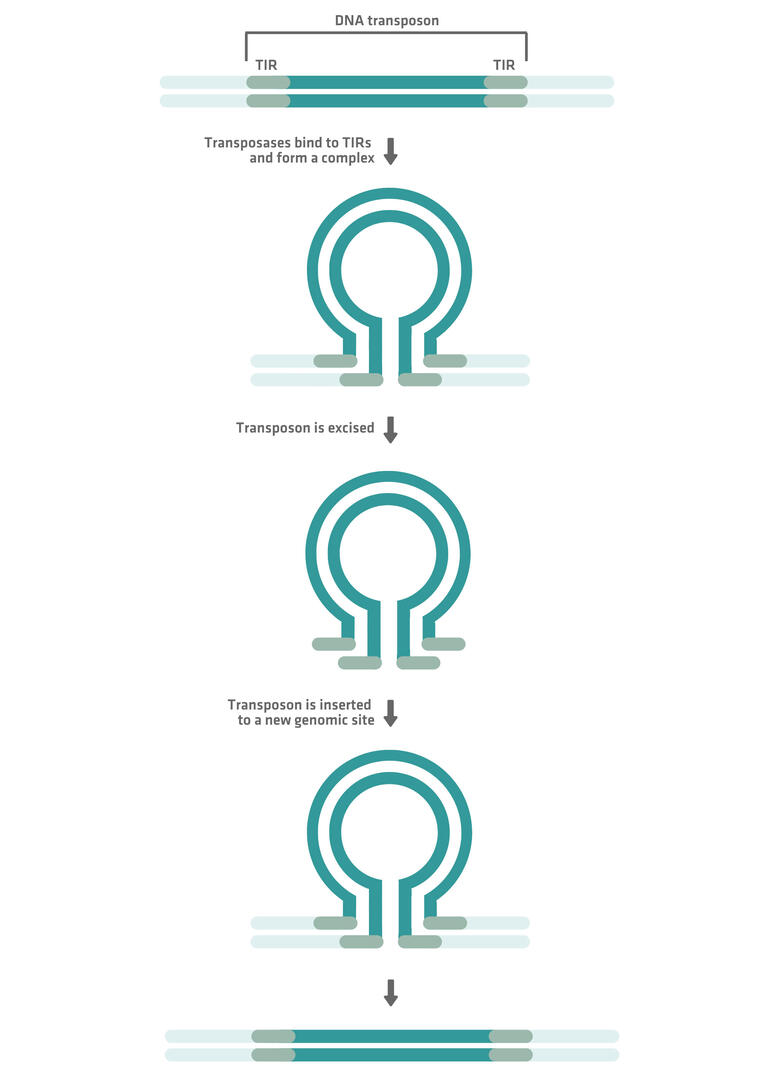
DNA transposons: class II TEs
DNA transposons, also known as class II TEs, move through a 'cut-and-paste' mechanism:
- First, transposase enzymes recognize specific terminal inverted repeat (TIR) sequences at both ends of the transposon.
- The transposases bind to these TIRs and form a complex.
- The entire transposon is excised from its original location.
- After excision, the transposases help insert the transposon into a new genomic site.
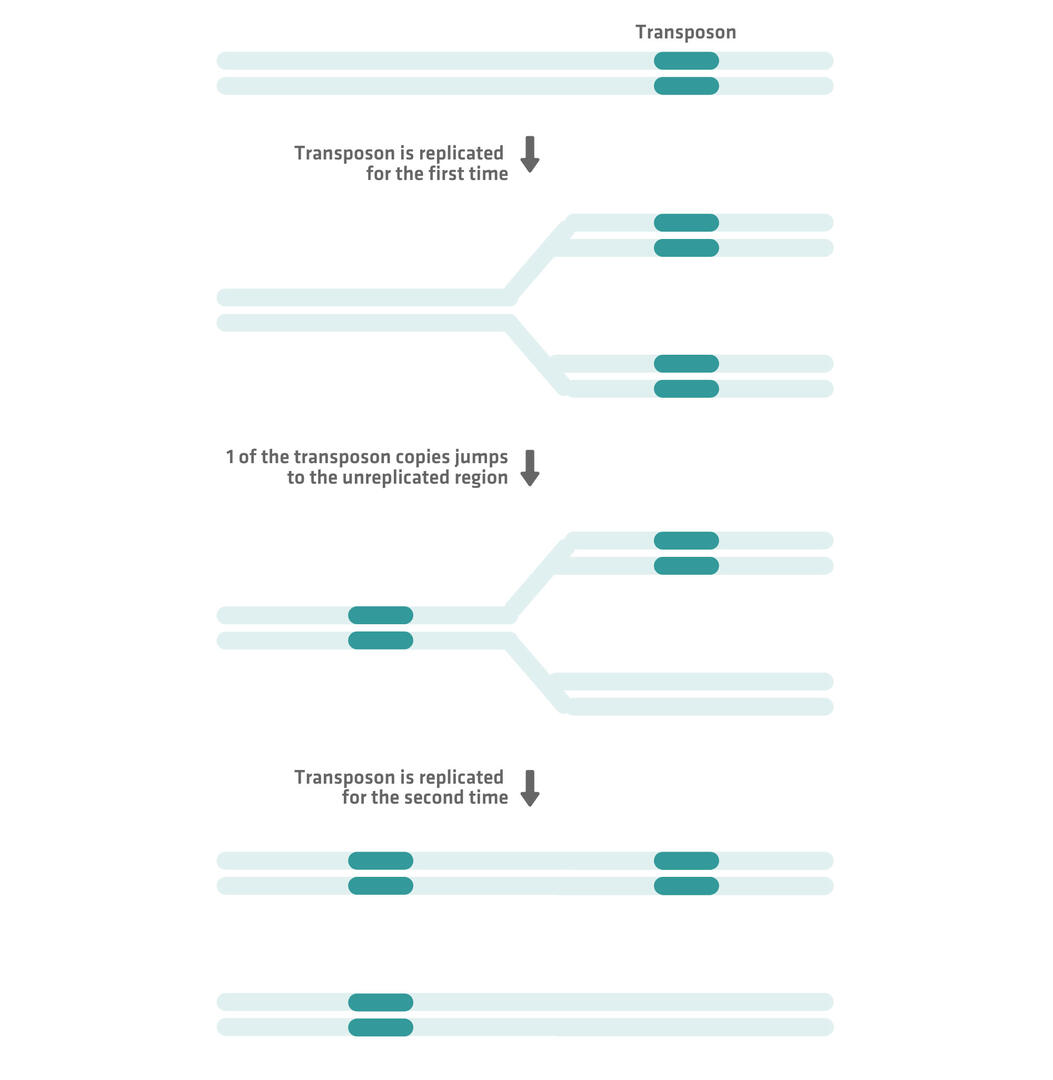
Despite using this 'cut-and-paste' mechanism, DNA transposons can still increase their copy number by moving during DNA replication. For example, if a DNA transposon moves from a region that has already been replicated to an unreplicated region of the genome, the replication process will ensure that 2 copies of the transposon are present in 1 of the resulting chromosomes:
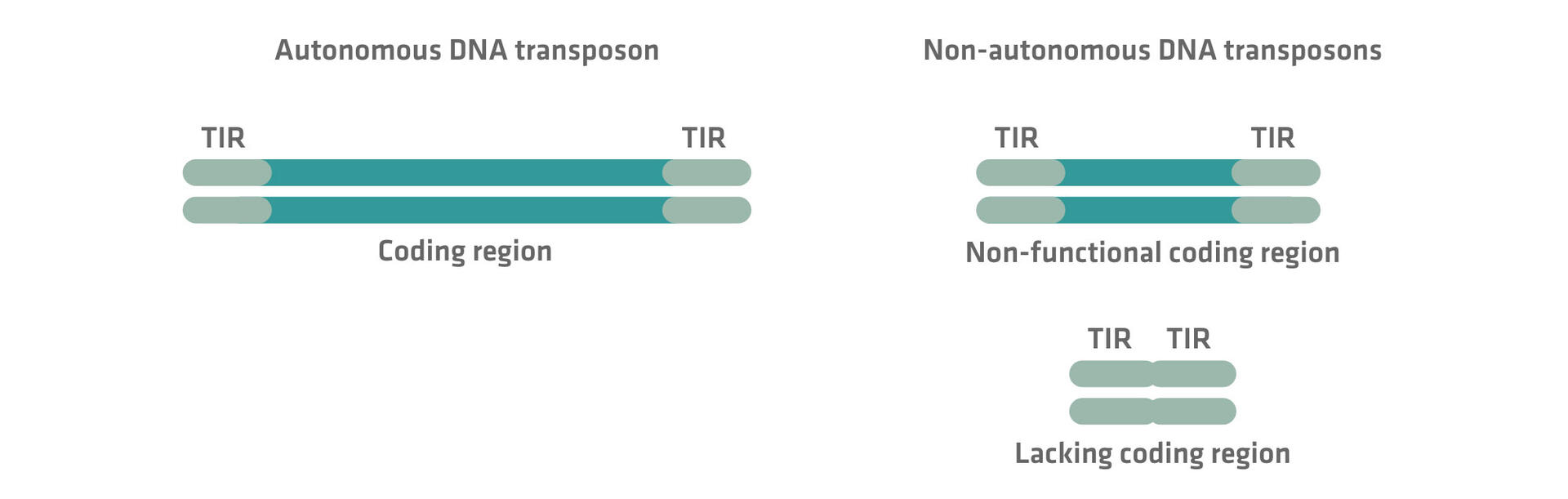
Autonomous vs non-autonomous transposons
The complexity of transposon classification is further increased by the distinction between autonomous and non-autonomous retrotransposons and DNA transposons.
As seen above, all transposons rely on proteins such as reverse transcriptases or transposases to move. Autonomous transposons contain all the genetic information needed to encode these components in their coding region. This enables them to autonomously transpose across the genome once activated. In comparison, non-autonomous transposons – which are usually shorter than their autonomous partners – lack some, or all, of the genetic information needed for transposition. However, they still contain all the necessary DNA sequences and motifs required to be recognized by the transposon machinery produced by other autonomous transposons, so they can still be transposed.
For example, autonomous DNA transposons feature a region coding for the transposase enzyme – essential for their ability to ‘jump around’ – flanked by 2 TIR sequences. In non-autonomous DNA transposons, the coding region is non-functional or missing, but the TIR sequences are intact, allowing them to be recognized and mobilized by a transposase enzyme encoded by an autonomous partner.
Similarly, autonomous LTR retrotransposons contain a region coding for reverse transcriptase and integrase enzymes between their LTR sequences, whereas this sequence is damaged or missing in non-autonomous forms.
In the case of non-LTR retrotransposons, autonomous versions are called long interspersed nuclear elements (LINEs), while non-autonomous versions are called short interspersed nuclear elements (SINEs).
Some non-autonomous transposons evolved from autonomous partners through deletions or mutations that disabled parts of the machinery needed for autonomous transposition. Others appear to have originated from non-coding genes – such as tRNAs or other genomic loci – which gained flanking regions recognizable by the transposon machinery. These versions have never encoded their own transposition proteins.
The first transposon discovered by Barbara McClintock in maize was the non-autonomous DNA transposon Ds (Dissociation). It requires an autonomous DNA transposon called Ac (Activator) that codes for the Ac transposase enzyme, in order to be excised and reintegrated into a new location in the genome.
Transposons and their impact on evolution and health
Transposons have a dual nature as both a threat to genome stability and a driver of genetic diversity. On the harmful side, they can inactivate genes or disrupt gene expression patterns by inserting into regulatory or coding regions, leading to diseases such as hemophilia, hypercholesterolemia and various cancers. However, transposons also play a key part in evolution. For instance, in plants, environmental stresses like climate change can trigger increased transposon activity, potentially giving rise to adaptive mutations that enhance survival.
To counter the potentially damaging effects of transposons, most organisms have evolved epigenetic mechanisms to regulate their activity. In humans, for example, regions of the genome rich in transposons are often tightly condensed, so that the transcription machinery cannot access them. This densely packed DNA is called heterochromatin, and it is formed through histone modifications (changes to the histone proteins around which the DNA is wound) and DNA methylation (the addition of methyl groups to cytosine bases). Other silencing mechanisms are based on RNAi or small RNAs, such as piRNAs, miRNAs or siRNAs, to specifically target and inactivate transposons.
Applications of transposons
Transposons can also be useful tools in the lab, with applications in mutagenesis, transgenesis and gene therapy.
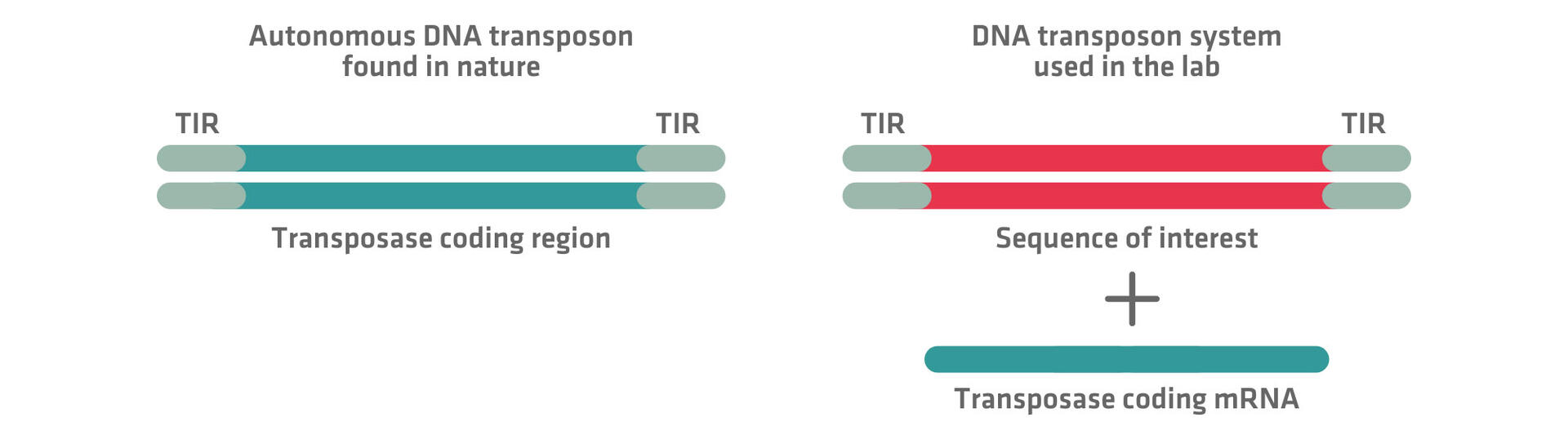
Researchers typically use a 2-component system based on naturally occurring autonomous DNA transposons for these applications. Unlike native transposons found in an organism's genome, these engineered systems replace the transposase coding region between the TIRs with a sequence of interest. As this component would be unable to move on its own, a transposase is provided as a second component, for example, in the form of an mRNA molecule encoding it. Some transposases randomly insert transposons in an unbiased fashion, whereas others are biased towards specific genetic features, such as AT-rich, GC-rich or active regions. This diversity provides researchers with a range of options tailored to their needs.
Transposon mutagenesis
Transposon mutagenesis is a technique allowing the study of gene function. Using DNA transposon systems, scientists introduce random mutations into an organism's genome. These mutations can then be located by identifying the insertion sites. Observing how these insertions affect the organism allows researchers to determine the roles of different genes.
Transposon transgenesis
Transposon mutagenesis usually aims to disrupt gene function to find out what a certain gene is responsible for, whereas transposon transgenesis focuses on introducing specific traits or functions into a target organism. The specific gene to be expressed is inserted into the target organism's genome using transposon systems. Researchers can apply this technique to cell lines, somatic cells or germ cells. Treating germ cells offers a distinct advantage: the transgene is transferred to all of the offspring. This approach can serve various purposes, including modeling human cancers in mice to test new treatments, or developing crops that are resistant to certain conditions.
Gene therapy
Transposon applications are also emerging in the field of human gene therapy, with transposon systems becoming more and more popular for delivering therapeutic genes into cells thanks to their advantages over traditional viral vectors:
- Their larger cargo capacity allows the delivery of longer DNA sequences
- They lack viral components, reducing the likelihood of inciting negative immune responses
- They offer easier and more cost-effective large scale production
- They provide a more predictable integration pattern
An example of a transposon-based gene therapy is CAR-T cells in the treatment of cancer. In contrast to normal T cells, CAR-T cells express a receptor specific to a protein on the surface of most malignant B cells.
Major transposon systems
The 3 most widely used transposon systems for lab applications are piggyBac (PB), Sleeping Beauty (SB) and Tol2 (from 'Transposable element of Oryzias latipes, number 2').
PiggyBac transposon system
The name of this DNA transposon system is linked to how it was discovered. When propagating baculovirus in a cell line derived from the cabbage looper moth, it was observed that mobile DNA elements from the insect cells could jump to the baculovirus genome, inspiring scientists to call it piggyBac.
The PB transposon system has a large cargo capacity of over 100 kb and can be used for genome engineering in a wide range of organisms, such as yeast, plants and mammals, including mice and humans.
An advantage of the PB transposon system is that the excision of PB is precise, usually leaving no trace behind at its original insertion site. This is important because leftover traces, or 'footprints', can sometimes cause unwanted changes in the DNA, such as mutations that can disrupt gene function. While this footprint-free movement is not crucial for most applications where the inserted genes are meant to stay put after integration, it becomes significant for workflows where a transposase enzyme remains active in the cell for a longer period, removing and integrating the PB transposon several times. Moreover, scientists have developed a version of PB transposase that can remove previously inserted sequences without reinserting them elsewhere, which is useful for temporary genetic modifications, such as creating induced pluripotent stem cells from adult tissues and then removing the reprogramming factors.
A further benefit of PB transposons is that they show a preference for integration near transcriptional start sites in the genome. This increases the chance that the insert ends up in an active region, either disrupting gene functions, which is the aim in mutagenesis applications, or being expressed, which is the aim in transgenesis and gene therapy applications.
Since its discovery, the PB transposon system has undergone multiple generations of enhancements, and hyperactive variants that integrate genes more efficiently have been developed.
Sleeping Beauty transposon system
SB is a synthetic DNA transposon that was 'awakened' by repairing inactivating mutations in ancient non-autonomous transposon copies discovered in various fish species. The reconstructed transposon can carry inserts of over 10 kb and functions in various vertebrates, including mice and humans, which is why it became a popular choice for genome engineering applications.
Since humans do not produce proteins similar to SB's transposase, the likelihood of an unintended transposition of the sequence of interest after its initial delivery and integration into human cells is very low. This property makes the SB transposon system an excellent candidate for gene therapy applications. However, unlike other transposon systems that favor integration near regulatory regions or within genes, SB integrates in a close-to-random manner, which means that more cells need to be treated to ensure expression of the inserted sequence.
Just like the PB system, SB has also undergone several modifications to improve its efficiency, and hyperactive versions – such as SB100X – are now available.
Tol2 transposon system
The Tol2 transposon system was discovered in 1996 in the Japanese medaka fish (Oryzias latipes), where it was identified due to an insertion causing an albino phenotype. It was the first DNA transposon found to be active in vertebrates. It can function in a variety of organisms – such as frogs, chicken, mice and humans – and is the preferred transposon system for mutagenesis and transgenesis applications in zebrafish.
Tol2 often integrates near transcription start sites and regulatory regions, which means that its chances of ending up in an active region are good. Moreover, it offers a cargo capacity of up to 11 kb and the transgenic sequences it inserts are reliably expressed without being silenced by host organisms.
Various improved versions of Tol2 are available, including a miniTol2 version that is shorter and shows an increased transposition activity.
Alternative transposon systems
The list of transposon systems described in this blog is not exhaustive. For instance, bacterial transposon systems like Tn5, Tn7 and Tn10 have also been adapted as molecular biology tools. Among these, Tn5 is predominantly used in NGS applications, Tn7 for gene integration into bacterial genomes, and Tn10 for mutagenesis and transgenesis applications.
Conclusion
The discovery of transposons revolutionized our understanding of genetics, revealing the dynamic nature of genomes. While these mobile elements can cause genetic disruptions and contribute to diseases, they also drive evolution and can be harnessed for applications in genetic research and gene therapy. Transposons are no longer just a fascinating genetic oddity – they are central players in the future of science and medicine.
Did you like this article?
Subscribe to our blog for more!
Ask our expert. Leave a comment!
Write us if you have any questions regarding the blog article.